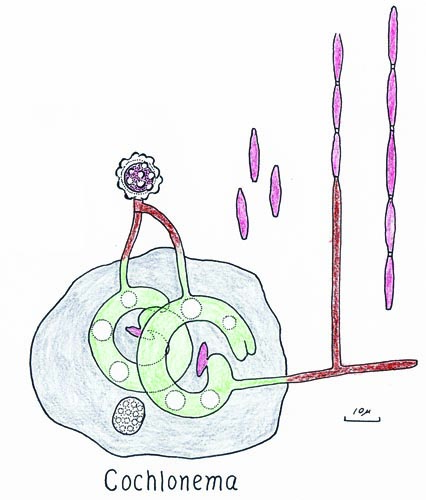
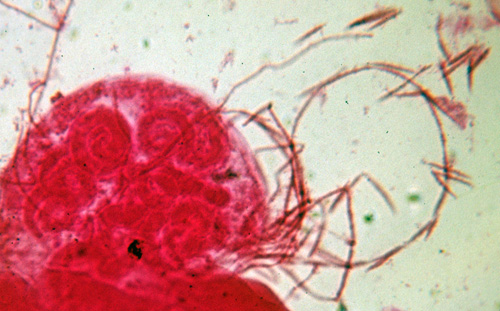
Cochlonema
COCHLONEMA Drechsler, 1935 (Mycologia 27: 18), 18 species (Drechsler, 1935).
Endoparasites of amoebae and rhizopods. Thallus coiled, rarely digitate, usually 3 or more times wider than the asexual hyphae; haustoria not formed. Fertile hyphae not extensive, simple or branched, erect portions producing multispored merosporangia. Spores cylindrical to fusiform, smooth-walled or irregular in outline. Zygospores with a depressed, warted or peg-like ornaments; suspensors hyphoid or somewhat enlarged or more or less parallel or entwined.
Type species: C. verrucosum
Species of Cochlonema:
C. agamum Drechsler, 1946 (Mycologia 38:132) (Drechsler, 1946; Miura, 1972).
C. bactrosporum Drechsler, 1939 (Mycologia 31:142) (Drechsler, 1939a).
C. bactrosporum var. longius Drechsler, 1942 (Mycologia 34:293) (Drechsler, 1942).
C. calosperma Drechsler, 1951 (Mycologia 43:167) (Drechsler, 1951; Miura, 1972).
C. cerasphorum Drechsler, 1959 (Mycologia 51:815) (Drechsler, 1959).
C. cylindricum Drechsler, 1937 (Mycologia 29:247) (Drechsler, 1937; Miura, 1972).
C. dolichosporum Drechsler, 1935 (Mycologia 27:20) (Drechsler, 1935).
C. euryblastum Drechsler, 1942 (Mycologia 34:287) (Drechsler, 1942; Miura, 1972).
C. explicatum Drechsler, 1955 (Mycologia 47:375) (Drechsler, 1955).
C. fusisporum Drechsler, 1939 (Mycologia 31:405) (Drechsler, 1939b).
C. lineare Jones, 1962 (Transactions of the British Mycological Society 45:348) (Jones, 1962).
C. megalosomum Drechsler, 1939 (Mycologia 31:134) (Drechsler, 1939a; Dayal and Srivastava, 1979).
C. megaspirema Drechsler, 1937 (Mycologia 29:240) (Drechsler, 1937).
C. odontosperma Drechsler, 1937 (Mycologia 29:233) (Drechsler, 1937; Miura, 1972; Saikawa and Sato, 1991).
C. ozotum Drechsler, 1945 (Mycologia 37:15) (Drechsler, 1945).
C. pumilum Drechsler, 1939 (Mycologia 31:401) (Drechsler, 1939b, 1945—zygospores).
C. pygmum Jones, 1959 (Transactions of the British Mycological Society 42:81). (Jones, 1959; considered a synonym of C. verrucosum fide Miura, 1972).
C. symplocum Drechsler, 1941 (Mycologia 33:266) (Drechsler, 1941).
C. verrucosum Drechsler, 1935 (Mycologia 27:19) (Drechsler, 1935; Miura, 1972).
Species of Cochlonema have been noted in only one plate or in several plates (Drechsler, 1935, 1937). Cochlonema spp. are parasites of amoebae and rhizopods. Species of Cochlonema are characterized by the formation of coiled thalli produced in the host and multispored merosporangia; haustoria are not formed. Zygospores are typical for these zoopagaceous fungi although zygosporangial ornamentation and suspensor morphology and arrangement are variable. One species, C. explicatum, that is a parasite of rhizopods, produces a branched thallus that is not coiled (Drechsler, 1955). In most species of Cochlonema, the spore is emptied of cytoplasmic contents but in C. explicatum (Drechsler, 1955) the spore remains filled with cytoplasm and is the origin of the fertile hyphae.
Merosporangia in Cochlonema are formed when the host stops moving. In C. megaspirema (Drechsler, 1937) the fertile hyphae arise from the coiled thallus, grow through the host pellicle and then the substrate. When the fertile hyphae become aerial they grow several millimeters high. Drechsler (1942) observed the white tufts of merosporangia of C. euryblastum by eye using lateral illumination 11 days after inoculation. In cultures of C. megalosomum (Drechsler, 1939a), the sporulating colonies are white specks to the naked eye but are a tuft of merosporangia when viewed with the microscope. When mature the merosporangia break apart leaving the spores scattered on the substrate surface. Spore ontogeny occurs by the formation of cytoplasm free zones at the end of each spore (Drechsler, 1937). Merosporangial formation occurs in abundance in many species and it is illustrated in C. agamum, C. bactrosporum, C. dolichosporum, C. euryblastum and C. verrucosum (Drechsler, 1935, 1939a, 1942, 1946). Many other species may produce a smaller number of spores, such as C. cylindricum and C. pumilum (Drechsler, 1939b, 1955). Spores of Cochlonema may be cylindrical, long-ellipsoid, or fusiform, and smooth or rough walled. Cylindrical spored species may be smooth-walled ([C. bactrosporum, C. bactrosporum var. longius, C. cylindricum, C. ozotum, C. pumilum; Drechsler, 1937, 1939a, 1939b, 1942, 1945) or rough-walled ([C. megaspirma, C. odontosperma; Drechsler, 1937). Another typical spore formed is fusiform that is smooth-walled ([C. fusisporum; Drechsler, 1939b) or rough-walled ([C. agamum, C. symplocum, C. verrucosum; Drechsler, 1935, 1941, 1946). Another interesting and unique spore, long-obovoid with an empty apex, is formed by C. dolichosporum (Drechsler, 1935). The other taxa produce ellipsoid or filaform to somewhat fiusiform spores with rough walls.
Drechsler (1941) noted that in Cochlonema symplocum zygospore formation did not occur until the heating quit working and then it started simultaneously. Drechsler (1939a) stated in C. megalospomum that only three infected amoebae produced zygospores of the hundreds that he observed; he thought that this taxon is heterothallic. Cochlonema agumum (Drechsler, 1935) forms azygospores on an apically distended suspensor. Sexual reproduction by the formation of globose zygospores has been reported in several species of Cochlonema. Internally formed sexual spores with a collapsed wall have been reported in C. bactrosporum and C. pumilum (Drechsler, 1939a, 1939b). Other species either have hemispherical ([C. megaspirema, C. symplocum, C. verrucosum; Drechsler, 1935, 1937, 1941) or columnar ornaments ([C. calosperma, C. megalosomum, C. odontosperma; Drechsler, 1937, 1939a, 1951) on zygospores formed outside the host. Suspensors are apposed and can be more or less parallel along the entire length and not swollen ([C. verrucosum; Drechlser, 1935) or apically inflated ([C. megalosomum, C. odontosperma; Drechsler, 1937, 1939a). Two species, C. megaspirema and C. symplocum (Drechsler, 1937, 1941) have suspensors that are wound around one another.
Bibliography
Dayal, R., and S.S. Srivastava. 1979. Three catenulate conidial Phycomycetes destructive to soil amoebae. Indian Phytopathology 32:409-412.
Drechsler, C. 1935. Some conidial Phycomycetes destructive to terricolous amoebae. Mycologia 27:6-40.
Drechsler, C. 1937. New Zoopagaceae destructive to soil rhizopods. Mycologia 29:229-249.
Drechsler, C. 1939a. A few new Zoöpagaceae destructive to large soil rhizopods. Mycologia 31:128-153.
Drechsler, C. 1939b. Five new Zoopagaceae destructive to rhizopods and nematodes. Mycologia 31:388-415.
Drechsler, C. 1941. Four Phycomycetes destructive to nematodes and rhizopods. Mycologia 33:248-269.
Drechsler, C. 1942. New species of Acaulopage and Cochlonema destructive to soil amoebae. Mycologia 34:274-297.
Drechsler, C. 1945. Several additional Phycomycetes subsisting on nematodes and amoebae. Mycologia 37:1-31.
Drechsler, C. 1946. Three new Zoöpagaceae subsisting on soil amoebae. Mycologia 38:120-143.
Drechsler, C. 1951. Various zoöpagaceous fungi subsisting on protozoans and eelworms. Mycologia 43:161-185.
Drechsler, C. 1955. Additional species of Zoopagaceae subsisting on rhizopods and eelworms. Mycologia 47:364-388.
Drechsler, C. 1959. Several Zoopagaceae subsisting on a nematode and some terricolous amoebae. Mycologia 51:787-823.
Jones, F.R. 1959. Some Zoopagales from Kenya. Transactions of the British Mycological Society 42:75-89.
Jones, F.R. 1962. New English Zoopagales. Transactions of the British Mycological Society 45:348-358.
Miura, K. 1972. Notes on filamentous fungi from Japan (9) The genus Cochlonema (Zoopagales). Journal of Japanese Botany. 47:204-214.
Saikawa, M., and H. Sato. 1991. Ultrastructure of Cochlonema odontosperma, an endoparasite in amoebae. Mycologia 83:403-408.
Updated Nov 05, 2007